Tutto ha inizio nelle gemme apicali: si chiamano così i germogli che stanno all’apice, in cima ai rametti di ogni pianta. Avrete sicuramente notato che nella maggior parte dei vegetali di taglia non proprio minimale, il germoglio che cresce alla fine del ramo, o più ancora quello che sta in cima a tutta la pianta, domina quelli sottostanti, crescendo molto più in fretta e determinando in questo modo la forma allungata del ramo e talvolta anche di tutta la pianta. Se così non fosse le piante crescerebbero in larghezza più che in altezza (ed in effetti alcune lo fanno). Le conifere come i pini e gli abeti rispettano molto bene la dominanza apicale, ma lo stesso fenomeno lo si può osservare un pò ovunque nel regno vegetale.
Nelle gemme apicali la pianta produce un ormone detto auxina, e chi ha dimestichezza con il greco capirà bene che il nome non è stato scelto a caso, dal momento che la radice “aux” sta per “crescita”. La cosa che non tutti immagineranno invece è che lo stesso ormone è implicato in due fenomeni apparentemente slegati l’uno dall’altro: la dominanza apicale ed il fatto che le piante si pieghino dalla parte della luce (fenomeno detto fototropismo).
Esistono auxine naturali e sintetiche. Quelle sintetiche sono prodotte industrialmente, hanno una struttura chimica diversa e vengono utilizzate per l’appunto per stimolare la crescita delle piante, a volte in modo così intenso da risultare per loro intollerabile (alcune sono infatti utilizzate come erbicidi), a volte per applicazioni diverse come ad esempio stimolare la formazione di radici nelle talee.
Ma focalizziamoci su quelle naturali, presenti in tutte le piante superiori. L’auxina più importante è sicuramente l’ acido 3-indolacetico, in sigla IAA.
La pianta lo produce a partire da un comune aminoacido, tra l’altro essenziale per l’uomo, il triptofano.
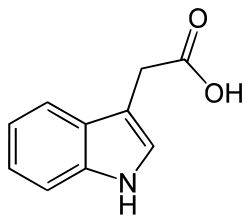
Fra le svariate azioni fisiologiche che l’IAA svolge nella pianta, 3 risultano essere particolarmente importanti per giustificarne l’azione:
1) determina la distensione delle cellule, quindi un notevole ingrossamento ed allungamento dei tessuti dove esso è presente in maggiore quantità ;
2) inibisce la crescita dei germogli
3) è fotosensibile, ovvero tende a degradarsi con la luce
Quando l’IAA è prodotto dalla gemma che sta in punta al ramo o a tutta la pianta (gemma apicale) esso si diffonde rapidamente con la linfa discendente e va nell’unica direzione dove può andare trovandosi all’estremità del ramo: centimetro per centimetro verso la base.
Qui in un primo tempo incontra i tessuti sottostanti la gemma che lo ha prodotto e, in funzione di quanto detto nel punto 1) favorisce il loro allungamento; successivamente incontra i germogli che stanno subito dietro alla gemma apicale, diciamo nei primi centimetri del ramo sotto la punta e qui, in funzione di quanto descritto al punto 2), ne inibisce la crescita.
Mentre scende lungo il ramo o la base della pianta esso si “consuma”, in buona parte anche per via della sua fotosensibilità, e questa è la ragione per la quale i germogli molto lontani dalla punta del ramo, diciamo quelli alla base, essendo meno inibiti tendono a crescere di più, dando vita a quella che a volte sembra essere una forma di crescita “a triangolo” del ramo.
La proprietà 3) invece sta alla base del fenomeno per il quale le piante si piegano dalla parte della luce. Degradandosi maggiormente in relazione all’intensità luminosa, l’acido 3-indolacetico viene a trovarsi in concentrazioni inferiori sul lato illuminato dei fusticini in accrescimento ed in concentrazioni maggiori nella parti in ombra: qui determina un maggiore rilassamento e quindi un ingrossamento delle cellule; come è facilmente intuibile, se un lato del fusto cresce più dell’altro, in ramo “si piegherà” in direzione del lato che cresce meno, in questo caso quello più illuminato, e si avrà la percezione che il fusto si pieghi dalla parte della luce.
La trattazione finora condotta è a dire il vero fondamentalmente biologica, o più specificamente fisiologica. Stiamo parlando di fisiologia vegetale.
Un chimico che tratti i fenomeni chimici all’interno di un organismo vivente, ovvero un biochimico, dovrebbe se non altro porsi alcuni quesiti più specifici.
Ad esempio iniziare a domandarsi “perché” o “in che modo” le auxine come l’IAA determinino una distensione della cellula vegetale, e cercherà di darsi una risposta utilizzando gli strumenti ed i metodi a sua disposizione.
In altre parole quello che le persone più petulanti chiamano “il perché del perché”. E la lista dei perché dei perché dei perché potrebbe crescere in linea di principio indefinitamente, facendoci passare da una sfera della conoscenza ad un’altra, sempre più interna, intima, basilare.
Classico ad esempio è il passaggio da una osservazione agronomica (es. le piante si piegano dalla parte della luce), ad una biologica (azione dell’auxina), ad una citologica-istologica ovvero relativa a tessuti e cellule (l’auxina determina un maggiore ingresso di acqua nella cellula, che pertanto di distende ed accresce le sue dimensioni), ad una biochimica (come fa l’auxina a fare entrare più acqua nella cellula?), ad una magari chimica in senso stretto, ovvero espressa in termini di semplici interazioni fra molecole organiche per terminare probabilmente in una spiegazione fisica dei fenomeni basilari che stanno dietro alle stesse interazioni chimico-organiche individuate.
E’ ovvio che, non essendo più ai tempi dello scienziato tuttologo e generalista, come ad esempio il rinascimentale Leonardo Da Vinci, ed essendosi il campo della conoscenza allargato in modo impressionante, non è più possibile trovare uno scienziato che riesca a seguire in modo approfondito tutti i livelli della conoscenza che possono spiegare i fenomeni, in questo caso dall’evidenza agronomica alle basi fisico-matematiche dei fenomeni. Relativamente alle auxine, alla base dell’ingrossamento delle cellule sembrerebbe esserci un aumento del volume dei vacuoli, che possono essere visti un po’ come le cisterne riempibili delle cellule vegetali: una cellula a vacuoli ben pieni può aumentare il suo volume anche di diverse volte rispetto ad una a vacuoli “scarichi”, sempre che la parete e la membrana cellulare riescano a dilatarsi in proporzione (un po’ come le pelle per una persona che sta ingrassando troppo in fretta).
Il materiale principale che riempie i vacuoli è semplicemente l’acqua. Le auxine determinano un incremento di ingresso dell’acqua nei vacuoli. L’argomento diventa più complicato se iniziamo a domandarci come fanno le auxine ad incrementare questo ingresso. Per semplificare si può comunque dire che in mezzo ci sta un’azione a carico della membrana cellulare, il “sacchetto” semipermeabile che racchiude oltre ai vacuoli tutti gli organuli propri della cellula: il passaggio di IAA da una cellula all’altra, solitamente nel senso discendente, dall’alto verso il basso, determina un flusso di protoni H+ (ovvero di idrogeno in forma ionica) da dentro la cellula a fuori. Questi determinano un’acidificazione dell’ambiente acquoso esterno alla cellula (pH 5.5 contro pH 7 dell’interno). Scaviamo ancora? Ad esempio chiediamoci cosa c’entra l’acidificazione dell’ambiente extracellulare con l’ingrossamento delle cellule. Scopriremo che nell’ambiente acido si attivano delle proteine presenti nella parete cellulare detta espansine (nome alquanto eloquente!): queste espansine provocano a loro volta lo slittamento delle micro-fibre di cellulosa che costituiscono la parere cellulare dei vegetali, considerata solitamente rigida, determinando un ammorbidimento della stessa. La parete cellulare meno rigida quindi sarà più suscettibile alla pressione che riceve dall’interno turgido della cellula, che per tanto si espanderà ingrandendosi.
Abbiamo detto tutto? Ovviamente la catena dei perché non finisce neppure ora.
Andando a scavare ancora più in profondità si entra come quasi sempre in questi casi nel campo della biologia molecolare: l’effetto sulla membrana cellulare è mediato dall’attivazione e/o dall’inibizione dell’espressione di alcuni geni, per altro di tipo piuttosto “generalista” in quanto rientrano a far parte, con variazioni tutto sommato minimali, del DNA della maggior parte delle piante.
La nota conclusiva segna un breve ritorno all’argomento iniziale, ovvero alla molteplicità delle diverse azioni che le auxine possono svolgere nelle piante.
Fra quelle più eclatanti dal un punto di vista fisiologico ma anche applicativo bisogna aggiungere che questi ormoni svolgono anche una funzione nel regolare la crescita della parte ipogea, ovvero sotterranea della pianta, in altre parole delle radici, regolando anche in questo caso la distensione cellulare e in ultima analisi anche qui la dominanza apicale. Da alcune decine di anni i coltivatori ma anche gli appassionati di giardinaggio ricorrono all’aiuto di preparati in polvere, più raramente in pasta o liquidi, contenenti auxine sintetiche per stimolare la formazione di radici sulle talee. Quando si taglia un ramo di una pianta, prima di interrarne una parte nell’intento di creare un individuo nuovo e completo, si è soliti cospargere la parte che sarà interrata con preparati a base soprattutto di acido naftalenacetico (NAA): a basse concentrazioni questa molecola, più resistente nel tempo del suo omologo “naturale” acido 3-indolacetico, stimola la generazione di nuove radici, ma a concentrazioni maggiori ne inibisce la crescita (per cui attenti ai dosaggi e soprattutto a non cospargere radici già formate!).
Ancora una volta un insegnamento morale che proviene dalla natura: il troppo storpia